6.6 Structure and Function of Blood Vessels
Learning Objectives
By the end of this section, you will be able to:
- Compare and contrast the three tunics that make up the walls of most blood vessels
- Distinguish between elastic arteries, muscular arteries, and arterioles on the basis of structure, location, and function
- Describe the basic structure of a capillary bed, from the supplying metarteriole to the venule into which it drains
- Explain the structure and function of venous valves in the large veins of the extremities
Blood is carried through the body via blood vessels. An artery is a blood vessel that carries blood away from the heart, where it branches into ever-smaller vessels. Eventually, the smallest arteries, vessels called arterioles, further branch into tiny capillaries, where nutrients and wastes are exchanged, and then combine with other vessels that exit capillaries to form venules, small blood vessels that carry blood to a vein, a larger blood vessel that returns blood to the heart.
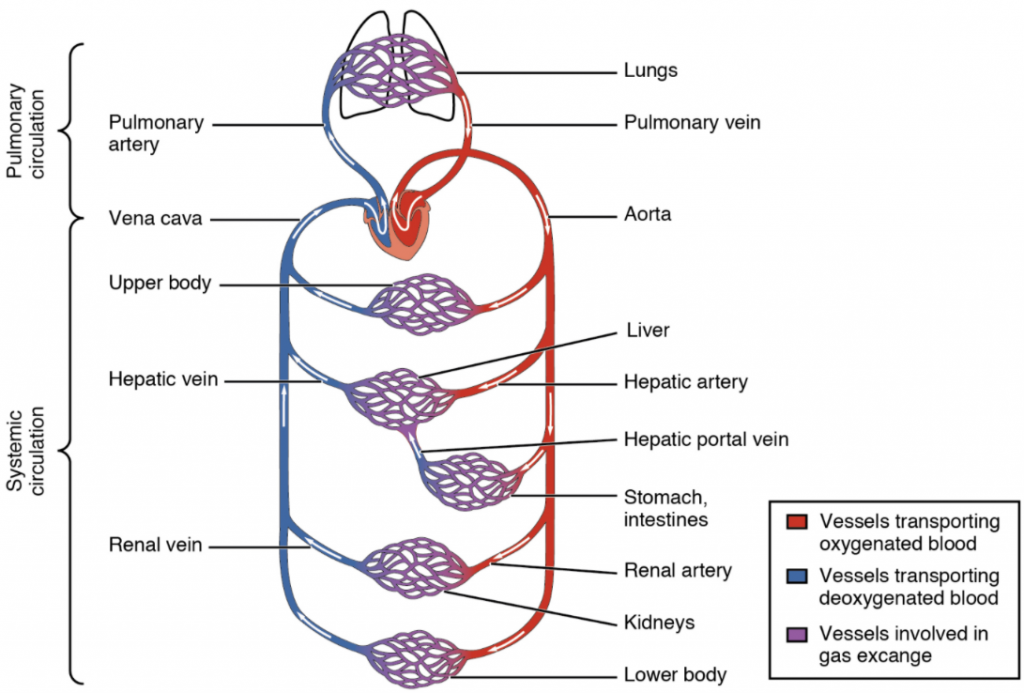
Arteries and veins transport blood in two distinct circuits: the systemic circuit and the pulmonary circuit (Figure 6.6.1). Systemic arteries provide blood rich in oxygen to the body’s tissues. The blood returned to the heart through systemic veins has less oxygen, since much of the oxygen carried by the arteries has been delivered to the cells. In contrast, in the pulmonary circuit, arteries carry blood low in oxygen exclusively to the lungs for gas exchange. Pulmonary veins then return freshly oxygenated blood from the lungs to the heart to be pumped back out into systemic circulation. Although arteries and veins differ structurally and functionally, they share certain features.
Shared Structures
Different types of blood vessels vary slightly in their structures, but they share the same general features. Arteries and arterioles have thicker walls than veins and venules because they are closer to the heart and receive blood that is surging at a far greater pressure (Figure 6.6.2). Each type of vessel has a lumen—a hollow passageway through which blood flows. Arteries have smaller lumens than veins, a characteristic that helps to maintain the pressure of blood moving through the system. Together, their thicker walls and smaller diameters give arterial lumens a more rounded appearance in cross section than the lumens of veins.
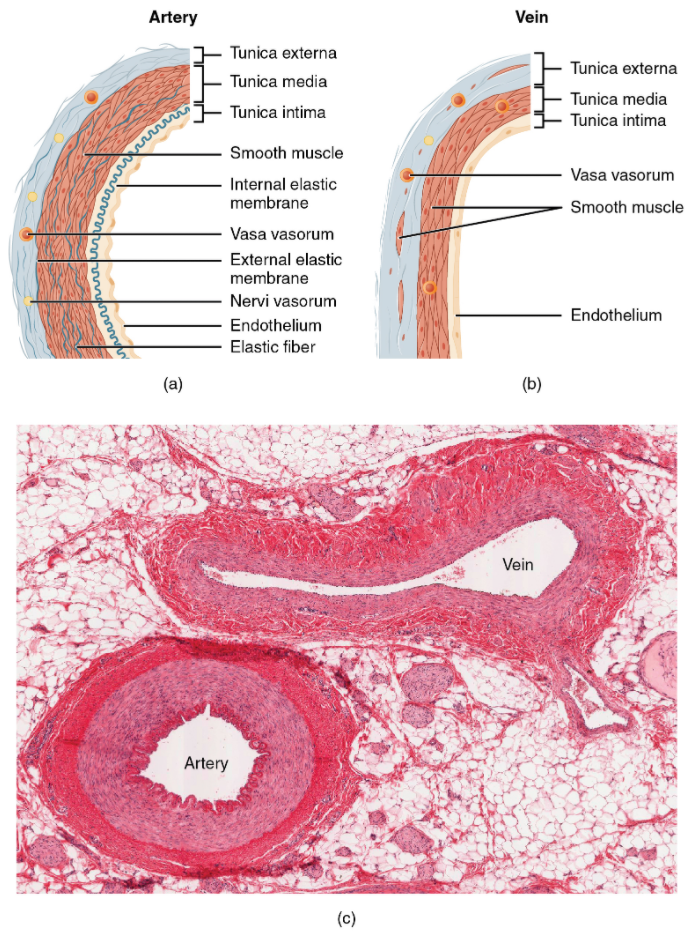
By the time blood has passed through capillaries and entered venules, the pressure initially exerted upon it by heart contractions has diminished. In other words, in comparison to arteries, venules and veins withstand a much lower pressure from the blood that flows through them. Their walls are considerably thinner and their lumens are correspondingly larger in diameter, allowing more blood to flow with less vessel resistance. In addition, many veins of the body, particularly those of the limbs, contain valves that assist the unidirectional flow of blood toward the heart. This is critical because blood flow becomes sluggish in the extremities, as a result of the lower pressure and the effects of gravity.
The walls of arteries and veins are largely composed of living cells and their products (including collagenous and elastic fibres); the cells require nourishment and produce waste. Since blood passes through the larger vessels relatively quickly, there is limited opportunity for blood in the lumen of the vessel to provide nourishment to or remove waste from the vessel’s cells. Further, the walls of the larger vessels are too thick for nutrients to diffuse through to all of the cells. Larger arteries and veins contain small blood vessels within their walls known as the vasa vasorum—literally “vessels of the vessel”—to provide them with this critical exchange. Since the pressure within arteries is relatively high, the vasa vasorum must function in the outer layers of the vessel (see Figure 6.2.2) or the pressure exerted by the blood passing through the vessel would collapse it, preventing any exchange from occurring. The lower pressure within veins allows the vasa vasorum to be located closer to the lumen. The restriction of the vasa vasorum to the outer layers of arteries is thought to be one reason that arterial diseases are more common than venous diseases, since its location makes it more difficult to nourish the cells of the arteries and remove waste products. There are also minute nerves within the walls of both types of vessels that control the contraction and dilation of smooth muscle. These minute nerves are known as the nervi vasorum.
Both arteries and veins have the same three distinct tissue layers, called tunics (from the Latin term tunica), for the garments first worn by ancient Romans; the term tunic is also used for some modern garments. From the most interior layer to the outer, these tunics are the tunica intima, the tunica media, and the tunica externa (see Figure 6.6.2). Table 6.6.1 compares and contrasts the tunics of the arteries and veins.
Table 6.6.1. Comparison of tunics in arteries and veins
| Arteries | Veins | |
| General appearance | Thick walls with small lumens | Thin walls with large lumens |
| Generally appear rounded | Generally appear flattened | |
| Tunica intima | Endothelium usually appears wavy due to constriction of smooth muscle | Endothelium appears smooth |
| Internal elastic membrane present in larger vessels | Internal elastic membrane absent | |
| Tunica media | Normally the thickest layer in arteries | Normally thinner than the tunica externa |
| Smooth muscle cells and elastic fibres predominate (the proportions of these vary with distance from the heart) | Smooth muscle cells and collagenous fibres predominate | |
| External elastic membrane present in larger vessels | Nervi vasorum and vasa vasorum present | |
| External elastic membrane absent | ||
| Tunica externa | Normally thinner than the tunica media in all but the largest arteries | Normally the thicket layer in veins |
| Collagenous and elastic fibres | Some smooth muscle fibres | |
| Nervi vasorum and vasa vasorum present | Nervi vasorum and vasa vasorum present |
Tunica Intima
The tunica intima (also called the tunica interna) is composed of epithelial and connective tissue layers. Lining the tunica intima is the specialised simple squamous epithelium called the endothelium, which is continuous throughout the entire vascular system, including the lining of the chambers of the heart. Damage to this endothelial lining and exposure of blood to the collagenous fibres beneath is one of the primary causes of clot formation. Until recently, the endothelium was viewed simply as the boundary between the blood in the lumen and the walls of the vessels. Recent studies, however, have shown that it is physiologically critical to such activities as helping to regulate capillary exchange and altering blood flow. The endothelium releases local chemicals called endothelins that can constrict the smooth muscle within the walls of the vessel to increase blood pressure. Uncompensated overproduction of endothelins may contribute to hypertension (high blood pressure) and cardiovascular disease.
Next to the endothelium is the basement membrane, or basal lamina, that effectively binds the endothelium to the connective tissue. The basement membrane provides strength while maintaining flexibility, and it is permeable, allowing materials to pass through it. The thin outer layer of the tunica intima contains a small amount of areolar connective tissue that consists primarily of elastic fibres to provide the vessel with additional flexibility; it also contains some collagenous fibres to provide additional strength.
In larger arteries, there is also a thick, distinct layer of elastic fibres known as the internal elastic membrane (also called the internal elastic lamina) at the boundary with the tunica media. Like the other components of the tunica intima, the internal elastic membrane provides structure while allowing the vessel to stretch. It is permeated with small openings that allow exchange of materials between the tunics. The internal elastic membrane is not apparent in veins. In addition, many veins, particularly in the lower limbs, contain valves formed by sections of thickened endothelium that are reinforced with connective tissue, extending into the lumen.
Under the microscope, the lumen and the entire tunica intima of a vein will appear smooth, whereas those of an artery will normally appear wavy because of the partial constriction of the smooth muscle in the tunica media, the next layer of blood vessel walls.
Tunica Media
The tunica media is the substantial middle layer of the vessel wall (see Figure 6.6.2). It is generally the thickest layer in arteries, and it is much thicker in arteries than it is in veins. The tunica media consists of layers of smooth muscle supported by connective tissue that is primarily made up of elastic fibres, most of which are arranged in circular sheets. Toward the outer portion of the tunic, there are also layers of longitudinal muscle. Contraction and relaxation of the circular muscles decrease and increase the diameter of the vessel lumen, respectively. Specifically, in arteries vasoconstriction decreases blood flow as the smooth muscle in the walls of the tunica media contracts, making the lumen narrower and increasing blood pressure. Similarly, vasodilation increases blood flow as the smooth muscle relaxes, allowing the lumen to widen and blood pressure to drop. Both vasoconstriction and vasodilation are regulated in part by small vascular nerves, known as nervi vasorum, or “nerves of the vessel,” that run within the walls of blood vessels. These are generally all sympathetic fibres, although some trigger vasodilation and others induce vasoconstriction, depending upon the nature of the neurotransmitter and receptors located on the target cell. Parasympathetic stimulation does trigger vasodilation as well as erection during sexual arousal in the external genitalia of both sexes. Nervous control over vessels tends to be more generalised than the specific targeting of individual blood vessels. Local controls, discussed later, account for this phenomenon. (Seek additional content for more information on these dynamic aspects of the autonomic nervous system.) Hormones and local chemicals also control blood vessels. Together, these neural and chemical mechanisms reduce or increase blood flow in response to changing body conditions, from exercise to hydration. Regulation of both blood flow and blood pressure is discussed in detail later in this chapter.
The smooth muscle layers of the tunica media are supported by a framework of collagenous fibres that also binds the tunica media to the inner and outer tunics. Along with the collagenous fibres are large numbers of elastic fibres that appear as wavy lines in prepared slides. Separating the tunica media from the outer tunica externa in larger arteries is the external elastic membrane (also called the external elastic lamina), which also appears wavy in slides. This structure is not usually seen in smaller arteries, nor is it seen in veins.
Tunica Externa
The outer tunic, the tunica externa (also called the tunica adventitia), is a substantial sheath of connective tissue composed primarily of collagenous fibres. Some bands of elastic fibres are found here as well. The tunica externa in veins also contains groups of smooth muscle fibres. This is normally the thickest tunic in veins and may be thicker than the tunica media in some larger arteries. The outer layers of the tunica externa are not distinct but rather blend with the surrounding connective tissue outside the vessel, helping to hold the vessel in relative position. If you are able to palpate some of the superficial veins on your upper limbs and try to move them, you will find that the tunica externa prevents this. If the tunica externa did not hold the vessel in place, any movement would likely result in disruption of blood flow.
Arteries
An artery is a blood vessel that conducts blood away from the heart. All arteries have relatively thick walls that can withstand the high pressure of blood ejected from the heart. However, those close to the heart have the thickest walls, containing a high percentage of elastic fibres in all three of their tunics. This type of artery is known as an elastic artery (Figure 6.6.3). Vessels larger than 10 mm in diameter are typically elastic. Their abundant elastic fibres allow them to expand, as blood pumped from the ventricles passes through them, and then to recoil after the surge has passed. If artery walls were rigid and unable to expand and recoil, their resistance to blood flow would greatly increase and blood pressure would rise to even higher levels, which would in turn require the heart to pump harder to increase the volume of blood expelled by each pump (the stroke volume) and maintain adequate pressure and flow. Artery walls would have to become even thicker in response to this increased pressure. The elastic recoil of the vascular wall helps to maintain the pressure gradient that drives the blood through the arterial system. An elastic artery is also known as a conducting artery, because the large diameter of the lumen enables it to accept a large volume of blood from the heart and conduct it to smaller branches.
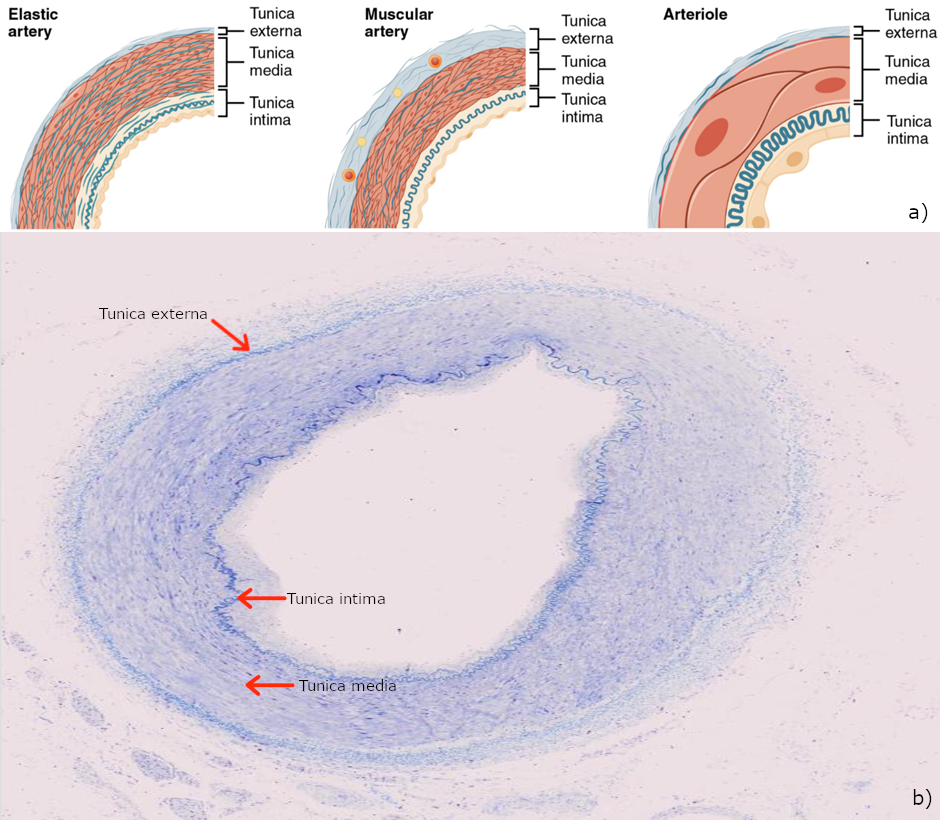
Farther from the heart, where the surge of blood has dampened, the percentage of elastic fibres in an artery’s tunica intima decreases and the amount of smooth muscle in its tunica media increases. The artery at this point is described as a muscular artery. The diameter of muscular arteries typically ranges from 0.1 mm to 10 mm. Their thick tunica media allows muscular arteries to play a leading role in vasoconstriction. In contrast, their decreased quantity of elastic fibres limits their ability to expand. Fortunately, because the blood pressure has eased by the time it reaches these more distant vessels, elasticity has become less important.
Notice that although the distinctions between elastic and muscular arteries are important, there is no “line of demarcation” where an elastic artery suddenly becomes muscular. Rather, there is a gradual transition as the vascular tree repeatedly branches. In turn, muscular arteries branch to distribute blood to the vast network of arterioles. For this reason, a muscular artery is also known as a distributing artery.
Arterioles
An arteriole is a very small artery that leads to a capillary. Arterioles have the same three tunics as the larger vessels, but the thickness of each is greatly diminished. The critical endothelial lining of the tunica intima is intact. The tunica media is restricted to one or two smooth muscle cell layers in thickness. The tunica externa remains but is very thin (see Figure 6.6.3).
With a lumen averaging 30 micrometres or less in diameter, arterioles are critical in slowing down—or resisting—blood flow and, thus, causing a substantial drop in blood pressure. Because of this, you may see them referred to as resistance vessels. The muscle fibres in arterioles are normally slightly contracted, causing arterioles to maintain a consistent muscle tone—in this case referred to as vascular tone—in a similar manner to the muscular tone of skeletal muscle. In reality, all blood vessels exhibit vascular tone due to the partial contraction of smooth muscle. The importance of the arterioles is that they will be the primary site of both resistance and regulation of blood pressure. The precise diameter of the lumen of an arteriole at any given moment is determined by neural and chemical controls, and vasoconstriction and vasodilation in the arterioles are the primary mechanisms for distribution of blood flow.
Capillaries
A capillary is a microscopic channel that supplies blood to the tissues themselves, a process called perfusion. Exchange of gases and other substances occurs in the capillaries between the blood and the surrounding cells and their tissue fluid (interstitial fluid). The diameter of a capillary lumen ranges from 5–10 micrometres; the smallest are just barely wide enough for an erythrocyte to squeeze through. Flow through capillaries is often described as microcirculation.
The wall of a capillary consists of the endothelial layer surrounded by a basement membrane with occasional smooth muscle fibres. There is some variation in wall structure: In a large capillary, several endothelial cells bordering each other may line the lumen; in a small capillary, there may be only a single cell layer that wraps around to contact itself.
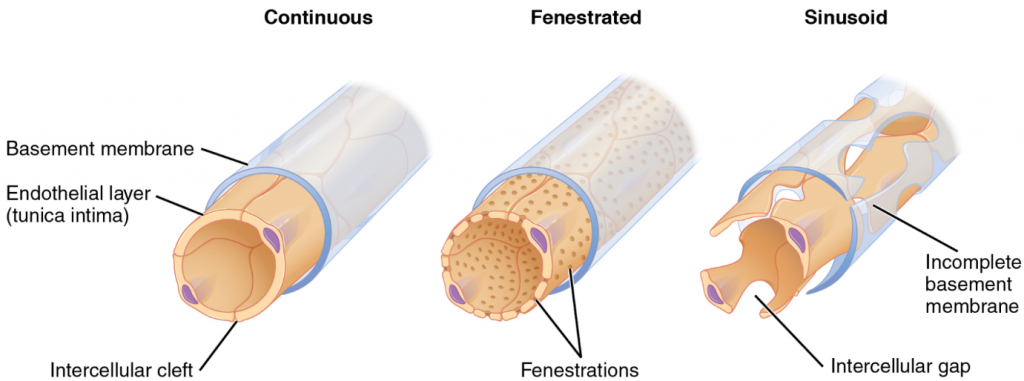
For capillaries to function, their walls must be leaky, allowing substances to pass through. There are three major types of capillaries, which differ according to their degree of “leakiness:” continuous, fenestrated, and sinusoid capillaries (Figure 6.6.4).
Continuous Capillaries
The most common type of capillary, the continuous capillary, is found in almost all vascularised tissues. Continuous capillaries are characterised by a complete endothelial lining with tight junctions between endothelial cells. Although a tight junction is usually impermeable and only allows for the passage of water and ions, they are often incomplete in capillaries, leaving intercellular clefts that allow for exchange of water and other very small molecules between the blood plasma and the interstitial fluid. Substances that can pass between cells include metabolic products, such as glucose, water, and small hydrophobic molecules like gases and hormones, as well as various leukocytes. Continuous capillaries not associated with the brain are rich in transport vesicles, contributing to either endocytosis or exocytosis. Those in the brain are part of the blood-brain barrier. Here, there are tight junctions and no intercellular clefts, plus a thick basement membrane and astrocyte extensions called end feet; these structures combine to prevent the movement of nearly all substances.
Fenestrated Capillaries
A fenestrated capillary is one that has pores (or fenestrations) in addition to tight junctions in the endothelial lining. These make the capillary permeable to larger molecules. The number of fenestrations and their degree of permeability vary, however, according to their location. Fenestrated capillaries are common in the small intestine, which is the primary site of nutrient absorption, as well as in the kidneys, which filter the blood. They are also found in the choroid plexus of the brain and many endocrine structures, including the hypothalamus, pituitary, pineal and thyroid glands.
Sinusoid Capillaries
A sinusoid capillary (or sinusoid) is the least common type of capillary. Sinusoid capillaries are flattened, and they have extensive intercellular gaps and incomplete basement membranes, in addition to intercellular clefts and fenestrations. This gives them an appearance not unlike Swiss cheese. These very large openings allow for the passage of the largest molecules, including plasma proteins and even cells. Blood flow through sinusoids is very slow, allowing more time for exchange of gases, nutrients, and wastes. Sinusoids are found in the liver and spleen, bone marrow, lymph nodes (where they carry lymph, not blood), and many endocrine glands including the pituitary and adrenal glands. Without these specialised capillaries, these organs would not be able to provide their myriad of functions. For example, when bone marrow forms new blood cells, the cells must enter the blood supply and can only do so through the large openings of a sinusoid capillary; they cannot pass through the small openings of continuous or fenestrated capillaries. The liver also requires extensive specialised sinusoid capillaries in order to process the materials brought to it by the hepatic portal vein from both the digestive tract and spleen, and to release plasma proteins into circulation.
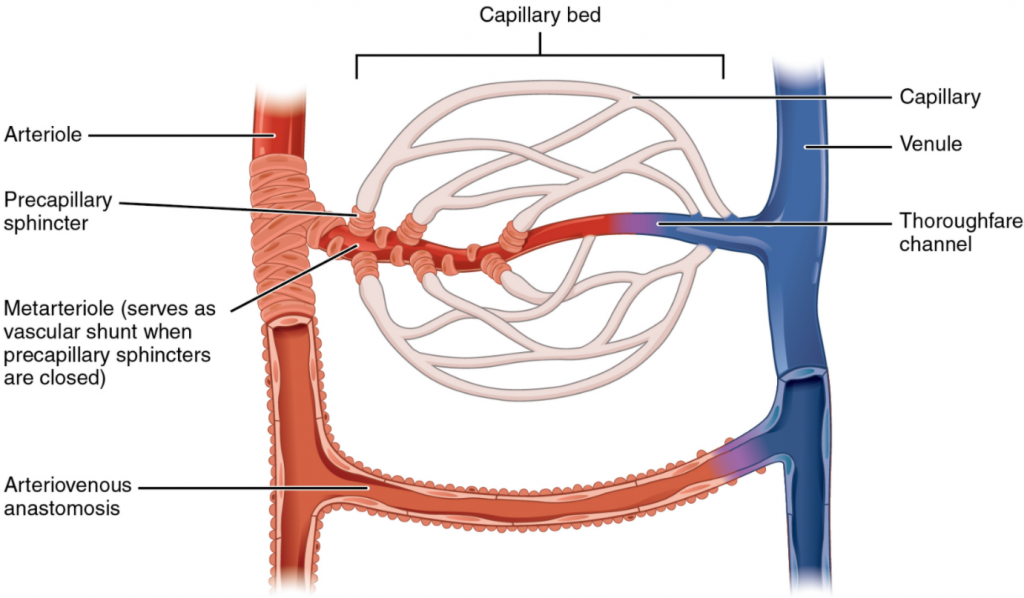
Metarterioles and Capillary Beds
A metarteriole is a type of vessel that has structural characteristics of both an arteriole and a capillary. Slightly larger than the typical capillary, the smooth muscle of the tunica media of the metarteriole is not continuous but forms rings of smooth muscle (sphincters) prior to the entrance to the capillaries. Each metarteriole arises from a terminal arteriole and branches to supply blood to a capillary bed that may consist of 10–100 capillaries.
The precapillary sphincters, circular smooth muscle cells that surround the capillary at its origin with the metarteriole, tightly regulate the flow of blood from a metarteriole to the capillaries it supplies. Their function is critical: If all of the capillary beds in the body were to open simultaneously, they would collectively hold every drop of blood in the body and there would be none in the arteries, arterioles, venules, veins, or the heart itself. Normally, the precapillary sphincters are closed. When the surrounding tissues need oxygen and have excess waste products, the precapillary sphincters open, allowing blood to flow through and exchange to occur before closing once more (Figure 6.6.5). If all of the precapillary sphincters in a capillary bed are closed, blood will flow from the metarteriole directly into a thoroughfare channel and then into the venous circulation, bypassing the capillary bed entirely. This creates what is known as a vascular shunt. In addition, an arteriovenous anastomosis may bypass the capillary bed and lead directly to the venous system.
Although you might expect blood flow through a capillary bed to be smooth, in reality, it moves with an irregular, pulsating flow. This pattern is called vasomotion and is regulated by chemical signals that are triggered in response to changes in internal conditions, such as oxygen, carbon dioxide, hydrogen ion, and lactic acid levels. For example, during strenuous exercise when oxygen levels decrease and carbon dioxide, hydrogen ion, and lactic acid levels all increase, the capillary beds in skeletal muscle are open, as they would be in the digestive system when nutrients are present in the digestive tract. During sleep or rest periods, vessels in both areas are largely closed; they open only occasionally to allow oxygen and nutrient supplies to travel to the tissues to maintain basic life processes.
Venules
A venule is an extremely small vein, generally 8–100 micrometres in diameter. Postcapillary venules join multiple capillaries exiting from a capillary bed. Multiple venules join to form veins. The walls of venules consist of endothelium, a thin middle layer with a few muscle cells and elastic fibres, plus an outer layer of connective tissue fibres that constitute a very thin tunica externa (Figure 6.6.6). Venules as well as capillaries are the primary sites of emigration or diapedesis, in which the white blood cells adhere to the endothelial lining of the vessels and then squeeze through adjacent cells to enter the tissue fluid.
Veins
A vein is a blood vessel that conducts blood toward the heart. Compared to arteries, veins are thin-walled vessels with large and irregular lumens (see Figure 6.6.6). Because they are low-pressure vessels, larger veins are commonly equipped with valves that promote the unidirectional flow of blood toward the heart and prevent backflow toward the capillaries caused by the inherent low blood pressure in veins as well as the pull of gravity. Table 2 compares the features of arteries and veins.
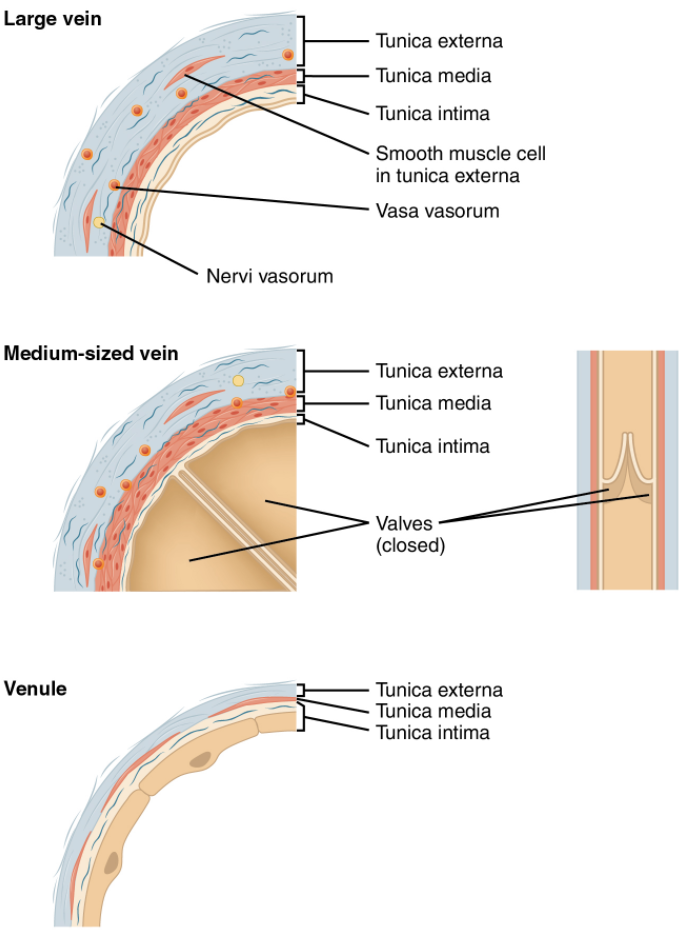
Table 6.6.2. Comparison of arteries and veins
| Arteries | Veins | |
| Direction of blood flow | Conducts blood away from heart | Conducts blood toward the heart |
| General appearance | Rounded | Irregular, often collapsed |
| Pressure | High | Low |
| Wall thickness | Thick | Thin |
| Relative oxygen concentration | Higher in systemic arteries
Lower in pulmonary arteries |
Lower in systemic veins
Higher in pulmonary veins |
| Valves | Not present | Present most commonly in limbs and in veins inferior to the heart |
Disorders of the Cardiovascular System: Oedema and Varicose Veins
Despite the presence of valves and the contributions of other anatomical and physiological adaptations we will cover shortly, over the course of a day, some blood will inevitably pool, especially in the lower limbs, due to the pull of gravity. Any blood that accumulates in a vein will increase the pressure within it, which can then be reflected back into the smaller veins, venules, and eventually even the capillaries. Increased pressure will promote the flow of fluids out of the capillaries and into the interstitial fluid. The presence of excess tissue fluid around the cells leads to a condition called oedema.
Most people experience a daily accumulation of tissue fluid, especially if they spend much of their work life on their feet (like most health professionals). However, clinical oedema goes beyond normal swelling and requires medical treatment. Oedema has many potential causes, including hypertension and heart failure, severe protein deficiency, renal failure, and many others. In order to treat oedema, which is a sign rather than a discrete disorder, the underlying cause must be diagnosed and alleviated.
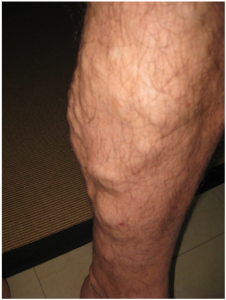
Oedema may be accompanied by varicose veins, especially in the superficial veins of the legs (Figure 6.6.7). This disorder arises when defective valves allow blood to accumulate within the veins, causing them to distend, twist, and become visible on the surface of the integument. Varicose veins may occur in both sexes but are more common in women and are often related to pregnancy. More than simple cosmetic blemishes, varicose veins are often painful and sometimes itchy or throbbing. Without treatment, they tend to grow worse over time. The use of support hose, as well as elevating the feet and legs whenever possible, may be helpful in alleviating this condition. Laser surgery and interventional radiologic procedures can reduce the size and severity of varicose veins. Severe cases may require conventional surgery to remove the damaged vessels. As there are typically redundant circulation patterns, that is, anastomoses, for the smaller and more superficial veins, removal does not typically impair the circulation. There is evidence that patients with varicose veins suffer a greater risk of developing a thrombus or clot.
Veins as Blood Reservoirs
In addition to their primary function of returning blood to the heart, veins may be considered blood reservoirs, since systemic veins contain approximately 64 percent of the blood volume at any given time (Figure 6.6.8). Their ability to hold this much blood is due to their high capacitance, that is, their capacity to distend (expand) readily to store a high volume of blood, even at a low pressure. The large lumens and relatively thin walls of veins make them far more distensible than arteries; thus, they are said to be capacitance vessels.
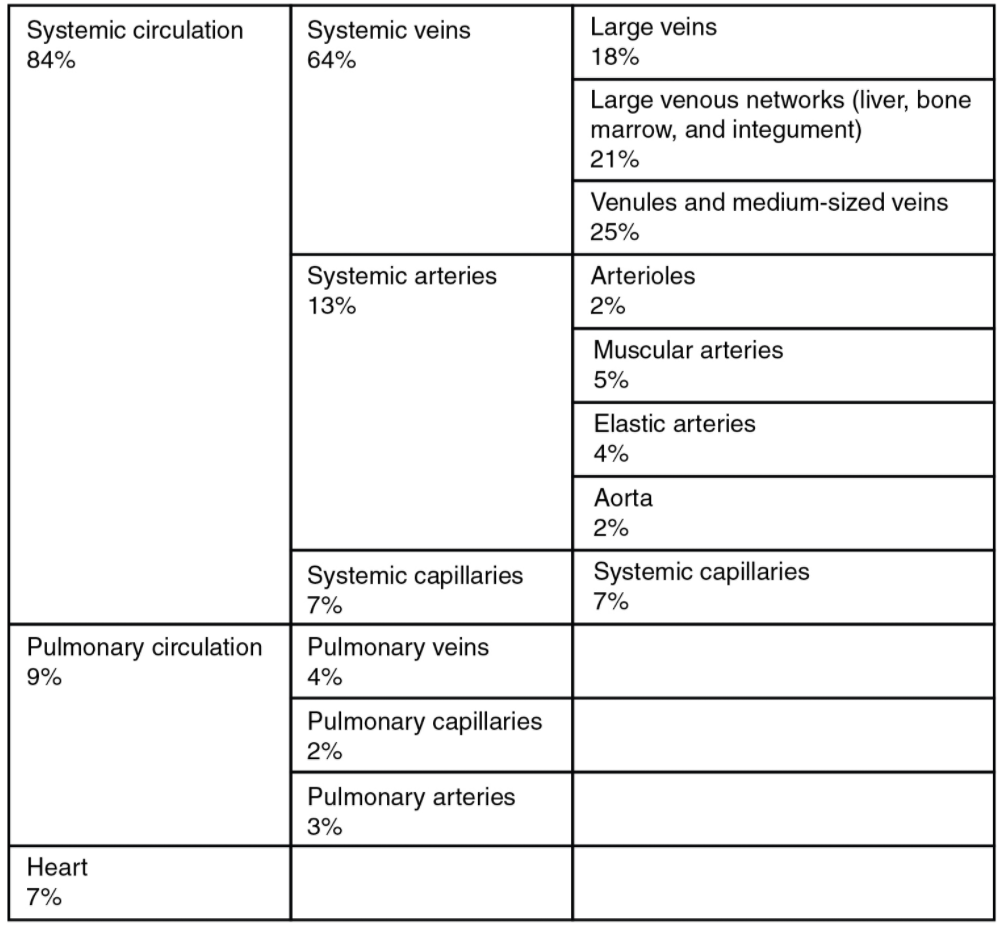
When blood flow needs to be redistributed to other portions of the body, the vasomotor centre located in the medulla oblongata sends sympathetic stimulation to the smooth muscles in the walls of the veins, causing constriction—or in this case, venoconstriction. Less dramatic than the vasoconstriction seen in smaller arteries and arterioles, venoconstriction may be likened to a “stiffening” of the vessel wall. This increases pressure on the blood within the veins, speeding its return to the heart. As you will note in Figure 6.6.8, approximately 21 percent of the venous blood is located in venous networks within the liver, bone marrow, and integument. This volume of blood is referred to as venous reserve. Through venoconstriction, this “reserve” volume of blood can get back to the heart more quickly for redistribution to other parts of the circulation.
Career Connection
Vascular Surgeons and Technicians
Vascular surgery is a specialty in which the physician deals primarily with diseases of the vascular portion of the cardiovascular system. This includes repair and replacement of diseased or damaged vessels, removal of plaque from vessels, minimally invasive procedures including the insertion of venous catheters, and traditional surgery. Following completion of medical school, the physician generally completes a five-year surgical residency followed by an additional one to two years of vascular specialty training. In Australia, most vascular surgeons are a member of the Vascular Surgeons in Australia and New Zealand (ANZSVS) which is the principal body for the education and training of vascular surgeons in Australia and New Zealand. This is similar to the Society of Vascular Surgery in the United States.
Vascular technicians are specialists in imaging technologies that provide information on the health of the vascular system. They may also assist physicians in treating disorders involving the arteries and veins. This profession often overlaps with cardiovascular technology, which would also include treatments involving the heart. In Australia, a cardiac surgeon may earn between $99k – $509k per annum.
Section Review
Blood pumped by the heart flows through a series of vessels known as arteries, arterioles, capillaries, venules, and veins before returning to the heart. Arteries transport blood away from the heart and branch into smaller vessels, forming arterioles. Arterioles distribute blood to capillary beds, the sites of exchange with the body tissues. Capillaries lead back to small vessels known as venules that flow into the larger veins and eventually back to the heart.
The arterial system is a relatively high-pressure system, so arteries have thick walls that appear round in cross section. The venous system is a lower-pressure system, containing veins that have larger lumens and thinner walls. They often appear flattened. Arteries, arterioles, venules, and veins are composed of three tunics known as the tunica intima, tunica media, and tunica externa. Capillaries have only a tunica intima layer. The tunica intima is a thin layer composed of a simple squamous epithelium known as endothelium and a small amount of connective tissue. The tunica media is a thicker area composed of variable amounts of smooth muscle and connective tissue. It is the thickest layer in all but the largest arteries. The tunica externa is primarily a layer of connective tissue, although in veins, it also contains some smooth muscle. Blood flow through vessels can be dramatically influenced by vasoconstriction and vasodilation in their walls.
Review Questions
Critical Thinking Questions
Click the drop down below to review the terms learned from this chapter.
